Ako môžu hliníkové adjuvanty vo vakcínach spôsobiť autizmus
alebo vakcíny poškodzujú nervový systém bábätiek
CDC (Centers for Disease Control and Prevention = Stredisko pre potláčanie a predchádzanie chorobám v USA; obdoba slovenského Úradu verejného zdravotníctva (ÚVZ SR) — pozn. prekl.) uvádza, že vakcíny a zložky vakcín boli vyvrátené ako možné príčiny autizmu. Tvrdenia CDC sú všeobecné a zahŕňajú všetky vakcíny a zložky vakcín. Napr. CDC tvrdí na svojej internetovej stránke (august 2017):
„Vakcíny nespôsobujú autizmus.“
„Neexistuje súvis medzi vakcínami a autizmom.“
„… žiadna súvislosť nebola zistená medzi akoukoľvek zložkou vakcín a poruchou autistického spektra.“
Tieto tvrdenia nepodporuje dostupná veda. Dôkazy, potvrdzujúce tieto tvrdenia CDC, sú obmedzené na MMR vakcínu (Taylor 2014), konzervant thimerosal (Taylor 2014) a pôsobeniu antigénu z vakcíny (DeStefano 2013).
Dr. Frank DeStefano z Oddelenia pre bezpečnosť vakcín CDC je spoluautorom štúdie (Glanz 2015), v ktorej sa uvádza:
„Dodnes nebola vypracovaná žiadna populačná štúdia, špecificky navrhnutá na vyhodnotenie spojitosti klinicky zmysluplných účinkov s neantigénnymi zložkami inými než thimerosal.“
Toto tvrdenie sa (okrem iných zložiek vakcín) týka aj hliníkového adjuvantu (pomocnej látky). Štúdie MMR vakcín (proti osýpkam, mumpsu a ružienke) nemôžu byť používané ako dôkaz o bezpečnosti iných vakcín, napr. vakcín, ktoré obsahujú hliník ako adjuvant. Zovšeobecňujúce tvrdenia, že žiadne vakcíny a žiadne ich zložky nespôsobujú autizmus, nemajú základ vo vedeckých dôkazoch. Tvrdenia CDC sú v skutočnosti v rozpore s obrovským, konzistentným a rastúcim množstvom vedeckých dôkazov, vrátane:
1. štúdií, preukazujúcich neurotoxické a neurozápalové účinky (napr. aktiváciu mikroglií) z dávok hliníkových adjuvantov nižších alebo približne rovnakých, ako sú dávky, ktoré dostávajú bábätká podľa očkovacieho kalendára CDC (Crepeaux 2017, Petrik 2007, Shaw 2013, Shaw 2009):
2. štúdie, spájajúce vakcíny s poškodeniami mozgu aktiváciou imunity (Zerbo 2016, Li 2015)
3. štúdie preukazujúce, že aktivácia imunity v skorom štádiu života je príčinným faktorom autizmu a ďalších neurovývinových porúch, ako i mentálnych ochorení ako napr. schizofrénia (Meyer 2009, Deverman 2009, Estes 2016, Kneusel 2014, Careaga 2017, Meyer 2014).
Hromadiace sa dôkazy naznačujú, že aktivácia imunity, podnietená vakcínou a špeciálne adjuvantom hliníka môže spôsobiť mentálne ochorenie a neurovývinové poruchy, vrátane autizmu.
V tejto analýze predstavíme vedecké dôkazy, že hliníkové adjuvanty môžu spôsobiť autizmus a ďalšie poškodenia mozgu. Zároveň vysvetlíme, prečo štúdie, údajne potvrdzujúce bezpečnosť hliníkových adjuvantov, nedokazujú bezpečnosť v prípade nežiaducich neurologických účinkov.
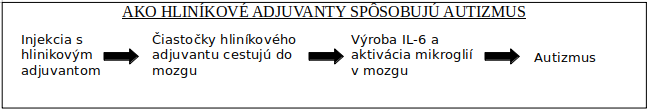
Aktivácia imunity: príčina autizmu a mentálnych ochorení
Výraz „aktivácia imunity“ opisuje aktiváciu bunkových zložiek imunitného systému. Vyvíjajúci sa mozog môže byť s celoživotnými dôsledkami poškodený aktiváciou imunity (Meyer 2009, Deverman 2009, Estes 2016, Kneusel 2014, Careaga 2017, Meyer 2014). Poškodenie aktiváciou imunity sa spája s autizmom, schizofréniou, depresiou a ďalšími mentálnymi ochoreniami alebo neurovývinovými poruchami. Vplyvy aktivácie imunity na mozog sú sprostredkované signalizačnými molekulami imunitného systému, obzvlášť cytokínmi (Estes 2016, Myer 2014, Smith 2007, Choi 2016, Pineda 2013)
Všeobecne sa akceptuje, že aktivácia imunity (napr. z prenosnej choroby) počas tehotenstva je rizikovým faktorom pre autizmus a schizofréniu u dieťaťa (Ciaranello 1995, Atladottir 2010, Brown 2012). Intenzita a priebeh aktivácie imunity a vylučovanie cytokínov sa javia byť dôležitým činiteľom ovplyvňujúcim riziko vzniku autizmu (Meyer 2014). Intenzívna aktivácia imunity sa spája s vyšším rizikom autizmu (Careaga 2017, Atladottir 2010). Chronický zápal sa spája s vyšším rizikom autizmu (Jones 2016, Zerbo 2014). Avšak nejestvuje dôkaz, že krátke trvanie a nízka intenzita aktivácie imunity, ktorá je následkom bežných detských ochorení, zvyšuje riziko autizmu. Načasovanie aktivácie imunity v súvislosti so stupňami vývoja mozgu je tiež dôležitým faktorom (Meyer 2006, Meyer 2009).
Experimenty na zvieratách testovali vplyvy aktivácie imunity počas tehotenstva a postnatálne na vývoj potomka (Meyer 2009, Deverman 2009, Estes 2016, Kenusel 2014, Careaga 2017, Meyer 2014). V týchto pokusoch je kotným zvieratám (myšiam, potkanom a opiciam) alebo ich potomkom po narodení vpichnutá neinfekčná imunnoaktivačná látka, ako napr. „poly-IC“ (ktorá napodobňuje vírusové ochorenie) alebo lipopolysacharid (LPS, ktorý napodobňuje bakteriálne ochorenie). Tieto látky spôsobujú aktiváciu imunitného systému bez nákazy. Vyvolajú horúčku a výrobu cytokínov a môžu mať podstatný vplyv na vývin mozgu, ak aktivácia je dostatočne intenzívna alebo predĺžená a ak je podstúpená počas zraniteľných vývinových etáp.
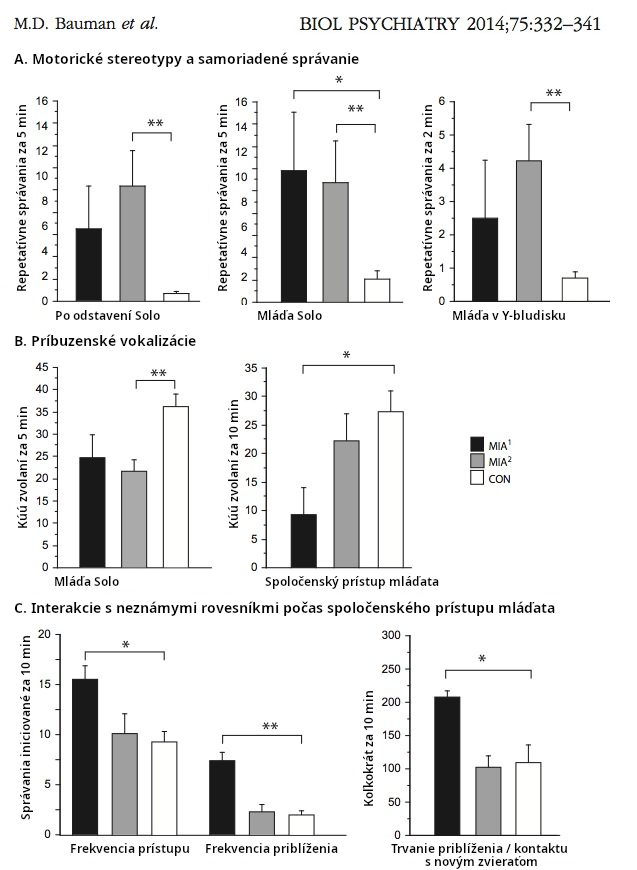
Aktivácia imunity, ako bolo preukázané na myšiach, spôsobuje tri kľúčové príznaky autistického správania sa: zníženú socializáciu a komunikáciu a zvýšenie repetitívneho správania (nezmyselné opakovanie nejakej činnosti - pozn. prekl.) (Malkova 2012). Dokázalo sa tiež, že aktivácia imunity spôsobuje poškodenie nervov (neuropatológiu, Weir 2015) a abnormality správania u opíc, ktoré pripomínajú správanie sa ľudských schizofrenikov a autistov (Bauman 2014, Machado 2015). Pozri obr. 2.
Aktivácia imunity tiež spôsobuje iné účinky (neprejavujúce sa v správaní sa) spájané s autizmom (tieto citácie spájajú aktiváciu imunity s týmito účinkami):
1. zníženie počtu Purkiněho buniek (Shi 2009)
2. mitochondriálna dysfunkcia (Giulivi 2013)
3. zvýšenie objemu mozgu (kvôli vplyvu IL-6, Wei 2012) a hustoty neurónov v mozgu (Smith 2012)
4. dlhohodobý chronický zápal mozgu (Garay 2012)
5. dysbióza (narušenie mikrobiómu, tzn. bakteriálnej mikroflóry (Hsiao 2013)
Tieto nebehaviorálne podobnosti (iné, než čo sa týka správania sa) ďalej len podporujú opodstatnenosť modelov aktivácie imunity pre ľudský autizmus. Nebehaviorálne (napr. fyziologické) účinky aktivácie imunity boli skúmané (Labouesse 2015).
Cytokínový interleukin-6 (IL-6) a interleukin-17a (IL-17) bol identifikovaný ako ten, ktorý sprostredkúva behaviorálne efekty aktivácie imunity(Smith 2007, Malkova 2012, Choi 2016, Pineda 2013, Wei 2012(a), Wei 2013, Parker-Athill 2010, Wei 2016). Zistenia o IL-6 boli znova potvrdené rôznymi výskumníkmi používajúcimi rôzne experimentálne metódy. Napr. v experimente s poly-IC, abnormálnemu správaniu je takmer úplne zabránené tým, že sa súčasne podá IL-6 blokujúca protilátka(Smith 2007, Pineda 2013). Injekcia IL-6 sama o seba zapríčiňuje abnormálne správanie, ktoré veľmi verne pripomína správanie, ktoré je výsledkom aktivácie imunity prostredníctvom poly-IC. (Smith 2007). Potlačenie IL-6 signalizácie v modeloch s genetickým autizmom (BTBR myši( normalizovalo sociálne a repetatívne správanie (Wei 2016). Tieto výsledky ukazujú, že IL-6 je zodpovedný za spôsobenie abnormálneho správania, ako je typické pre autizmus.
Pattersonovo laboratóriu v CalTech bolo prvé, ktoré prinieslo správu o tom, že IL-6 spôsobuje behaviorálne efekty, ako sú typické pre autizmus (Smith 2007). Dve štúdie tejto výskumnej skupiny tvrdia následovné:
„IL-6 je kľúčovým v procese, keď aktivácia imunity matky spôsobuje dlhodobé behaviorálne zmeny u potomka“. (Smith 2007)
„… blokovanie IL-6 predchádza viac ako 90% zmenám, ktoré sú pozorované u potomkov matiek, ktorým boli injekčne podané poly-IC, čo dokazuje, že zmeny genetickej expresie ako aj zmeny správania sú znormalizované eliminovaním IL-6 z materskej imúnnej reakcie“ (Smith 2007).
„IL-6 je nutný a dostatočný pre navodenie týchto efektov, nakoľko týmto efektom… u kotnej myši možno zabrániť injekčným podaním poly-IC a anti-IL-6 protilátky a možno ich napodobniť jedným injekčným podaním IL-6 matke.“ (Garay 2013)
Vystavenie mozgu zvýšenému IL-6 zostrojeným vírusom ukazuje, že vystavenie IL-6, započaté po narodení, spôsobilo správanie podobné autizmu (Wei 2012(a)). Táto štúdia Weiho tvrdí:
„Dokazujeme, že IL-6 je dôležitým mediátorom správania, typického pre autizmus. U myší so zvýšeným IL-6 sa v mozgu rozvinulo správanie, typické pre autizmu, vrátane poškodenia kognitívnej schopnosti, deficitu v učení, abnormálnych čŕt úzkostí a návykovosti, ako aj zníženú sociálnu interakciu inciovanú v neskorších štádiách. Tieto štúdie naznačujú, že zvýšenie IL-6 v mozgu môže modulovať isté patologické zmeny a prispievať k vývinu autizmu“ (Wei 2012(a)).
Nedávnejšie dôkazy ukazujú, že IL-17 pôsobí podobne ako IL-6, ktoré zapríčiňuje abnormality správania, typické pre autizmus, a atypické pre vývoj mozgovej kôry u myší (Choi 2016). Blokovanie buď IL-6 alebo IL-17 predchádza správaniu typickému pre autizmus, injekcia IL-17 sama o sebe spôsobuje správanie, typické pre autizmus (Choi 2016). Je známe, že IL-6 podnecuje IL-17 tým, že podporuje rozvoj Th17 buniek, ktoré produkujú IL-17.
Modely aktivácie imunity na zvieratách predstavujú platné modely pre neurologické (psychiatrické poruchy, vrátane autizmu (Estes 2016, Careaga 2017, Meyer 2014). Estes v recenzii z roku 2016 takto zdôvodňuje právoplatnosť týchto modelov aktivácie imunity pre ľudí:
„Tieto zvieracie modely MIA ( aktivácia imunity matky) spĺňajú všetky kritéria, požadované pre oprávnenosť modelu chorôb: Napodobňujú rizikový faktor príslušnýznámej chorobe (konštrukčná oprávnenosť(, ukazujú široké spektrum symptómov príslušnej choroby (priama pravoplatnosť( a môžu byť používané na predpovedanie efektívnosti liečby (prediktívna právoplatnosť).“ (Estes 2016)
Dôkazy naznačujú sprostredkujúcu rolu IL-6 a IL-17 v ľudskom autizme. Napr. IL-6 je podstatne zvýšené v mozočku v prípade ľudského autizmu (Wei 2011) a je vysoko zvýšené v niektorých oblastiach mozgu niektorých autistických jednotlivcov (Vargas 2005). Liečba ľudského autizmu s použitím protizápalového flavonoidového luteolínu zlepšuje autistické správanie u jednotlivcov, ktorých hladina IL-6 v krvi sa zároveň zníži. (Tsilioni 2015) Tento výsledok je konzistentný s príčinnou rolou IL-6 v ľudskom autizme. Aj IL-17 je zvýšené v prípade ľudského autizmu (Akintunde 2015, Al-Ayadhi 2012, Suzuki 2011). Vitamín D znižuje produkciu IL-17 (Bruce 2011, Wobke 2014, Drozdenko 2014) a zlepšuje autistické správanie u ľudí. (Saad 2016, Jia 2015). Zistenie o vitamíne D je konzistentné s príčinnou rolou IL-17 v ľudskom autizme.
Fungovanie IL-6 sa zdá byť podobné alebo identické u myší a ľudí. Žiadne rozdiely medzi myšami a ľuďmi vo fungovaní IL-6 neboli opísané v prehľade štúdií v roku 2004 (Mestas 2004).
Fungovanie IL-6 je priam zakonzervované naprieč druhmi (Brown 2014). Vývin centrálneho nervového systému u hlodavcov a ľudí je riadený tými istými princípmi (Brown 2014). Už aj preto fakt, že IL-6 zapríčiňuje u zvierat abnormálne správanie typické pre autizmus, si zasluhuje predpoklad právoplatnosti pre ľudí.
Aktivácia imunity je rizikovým faktorom pre autizmus, schizofréniu a ďalšie neurologické/psychiatrické poruchy. Cytokiny IL-6 a IL-17 sú zodpovedné za podnietenie behaviorálnych efektov aktivácie imunity, typických pre autizmus vo zvieracích modeloch. Dostupné dôkazové materiály podporujú príčinnú rolu IL-6 a IL-17 v ľudskom autizme.
Materská vs. Postnatálna aktivácia
Načasovanie aktivácie imunity je dôležitým faktorom ovplyvňujúcim dopad na mozog. Vyvíjajúci sa mozog je zraniteľný poškodením aktiváciou imunity. Dospelý, zrelý mozog nie je zjavne až tak zraniteľný. Citlivosť na aktiváciu i munitypravdepodobne klesá s tým, ako mozog dozrieva (Meyer 2014, Meyer 2007).
Vo väčšine experimentoch aktivácie imunity potomok bol vystavený aktivácii imunity počas gestácie (stimulovaním imunitného systému matky). Väčšina vakcín je však podávaná po narodení. Toto vzbudzuje otázku, či postnatálna aktivácia imunity môže mať podobné vplyvy na mozog ako aktivácia imunity matky. Rôznorodé dôkazy naznačujú, že mozog môže byť negatívne ovplyvnený postnatálne aktiváciou imunity. Experimenty s postnatálnou aktiváciou imunity, správy o ľudských prípadoch a zohľadnenie načasovania vývinu mozgu naznačujú, že ľudský mozog je zraniteľnejšína poškodenie aktiváciou imunity celé roky po narodení.
V experimentoch aktivácie imunity matky zápalové signalizovanie a niektoré cytokíny, napr. IL-6 prešli placentou do plodu. Následne, aktivácia imunity matky zapríčinila aktiváciu imunity a zvýšené cytokíny u plodu a v mozgu plodu (Oskvig 2012, Ghiani 2011).
Postnatálna aktivácia imunity môže mať negatívne neurologické vplyvy, vrátane zvýšenej náklonnosti k záchvatom (Chen 2013, Galic 2008), deficitom učenia a pamäte (Harre 2008) a zvýšenej tvorbe excitačných synapsií (Shen 2016). Záchvatové poruchy, dysfunkcia učenia a pamäte a zvýšené excitačné signalizovanie sú spájané s autizmom.
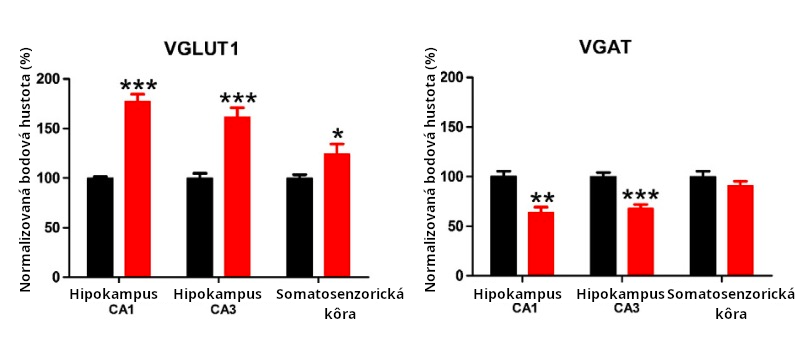
Zvýšené IL-6 v mozgu v postnatálnej dobe zapríčiňuje nerovnováhu nervového obvodu a podnecuje u myší správania typické pre autizmus (Wei 2012(a)). Obvodová nerovnováha pozorovaná v tejto štúdii Weiho bola excesom (nadbytkom) excitačných synapsií. Pozri obr. 3. Excesívne excitačné signalizovanie je pozorované v ľudskom autizme (Robertson 2016, Freyberg 2015). Fakticky nerovnováha medzi excitačným a inhibičným signalizovaním (naklonená k excitačnému) bola predpokladaná ako ústredná charakteristika autizmu (Roberston 2016, Freyberg 2015).
V experimente aktivácie materskej imunity u myší (Coiro 2015), autisticky relevantné správanie a abnormality dendritických tŕňov (relevantné pre autizmus a schizofréniu) boli zlepšené podaním protizápalových liekov postnatálne. Liek začali podávať pri narodení a pokračovali 2 týždne, čo zhruba zodpovedá veku 2 rokov u ľudí (Semple 2013). Tento výsledok naznačuje, že vývin mozgu je ovplyvnený postnatálnym zápalom, v čase zodpovedajúcom tomu, keď sú vakcíny podávané ľuďom.
Niekoľko prípadových štúdií opisuje predtým zdravé deti, že náhle sa u nich objavilo autistické správanie počas alebo následne po infekcii mozgu. Všetky prípady majú znaky intenzívneho mozgového zápalu. Tu sú stručné popisy:
Delong 1981: opisuje 3 deti vo veku 5,7 a 11 rokov s plne rozvinutým autistickým správaním spájaným s mozgovým zápalom. Zápal mozgu bol predpokladný v dvoch prípadoch a potvrdený v jednom. 5- a 7-roční plne vyzdraveli, 11-roční vyzdravel čiastočne.
Margues 2014: opisuje predtým zdravé 32-mesačné dievča, ktoré trpelo autistickou regresiou z virálnej infekcie nervového systému s pridruženým zápalom mozgu.
Ghaziuddin 2002: opisuje predtým zdravého 11-ročného chlapca, ktorý trpel trvalou autistickou regresiou po náhlej infekcii mozgu herpesom s pridruženým zápalom mozgu.
Gillberg 1986: opisuje predtým zdravé 14-ročné dievča s trvalou autistickou regresiou z infekcie mozgu herpesom s pridruženým zápalom mozgu.
Najskúpejšie vysvetlenie v týchto prípadoch je, že autistické správanie bolo výsledkom intenzívneho zápalu a produkcie citokínov v mozgu. Podobne tieto prípady naznačujú, že ľudský mozog počas detstva zostáva zraniteľnými na poškodenie aktiváciou imunity, kým táto zraniteľnosť sa takmer isto znižuje s dozrievaním. Chúlostivosť starších detí na autistické správanie spúšťané zápalom silno naznačuje, že mladšie deti vo veku 0-2 roky sú tiež zraniteľné. Nie je nerozumné tvrdiť, že niet dôkazov, ktoré by naznačovali, že vek 0-2 rokov (keď je podávaná väčšina vakcín), je jedinečne odolný voči poškodeniuaktiváciou imunity. Všetky dostupné dôkazy naznačujú opak.
Experimenty aktivácie imunity a prípadové štúdie sú konzistentné a naznačujú, že aktivácia imunity a zvýšené cytokíny v postnatálnom období môžu zapríčiniť poškodenie mozgu.
Ďalšia kritická otázka pre zváženie je, či vakcíny môžu zapríčiniť aktiváciu imunity a zvýšenie cytokínov v mozgu.
Postnatálne očkovanie ovplyvňuje vývin mozgu u zvieracích modelov
Prvá štúdia, ktorá testovala vplyv postnatálnej vakcinácie na vývin mozgu, bola publikovaná v roku 2015 (Li 2015). V tomto experimente, neonatálne potkany mali podanú vakcínu proti bacillus calmette-guerin (BCG) a proti hepatitíde B vakcínu (HBV) alebo kombináciu BCG+HBV načasovanú tak, aby imitovala očkovací kalendár ľudských detí. BCG a HBV vakcíny produkujú opačné efekty na mozog. Špeciálne BCG zlepšovala synaptickú plasticitu a dlhodobú potentiáciu (LTP, základ pre učenie a pamäť), HBV potláčala synaptickú plasticitu a LTP. BCG a HBV vakcíny tiež spôsobili opačné zmeny v niektorých úrovniach proteínov v synapsiách.
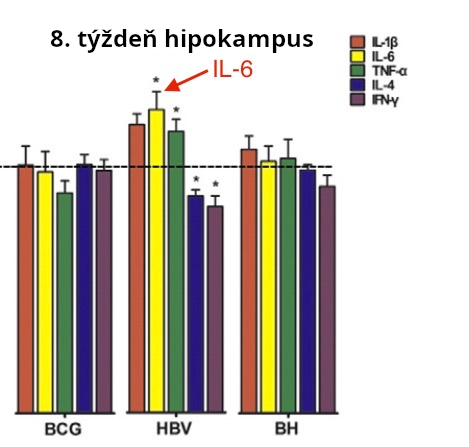
HBV vakcína (ale nie BCG vakcína) zvýšila genovú expresiu IL-6 v mozgu, zvýšená génová expresia pravdepodobne naznačuje zvýšenie mozgového IL-6. HBV vakcína obsahuje hliníkový adjuvant a BCG hliníkový adjuvant neobsahuje. Zjavne hliníkový adjuvant môže byť tou zložkou, ktorá je zodpovedná za zvýšenú genovú expresiu IL-6. Pozri obr. 4.
Li a spol ukázali, že vakcíny spôsobujú zmeny v mozgu vrátane 1) zmien v dlhodobej potenciácii (LTP - Hep B znížila LTP), 2) zmeny v dendritských tŕňoch a 3) zmeny v proteínovej expresii synapsií. Zmeny v synoptických proteínoch a dentrických tŕňoch boli pozorované v poruchách ľudských mozgov.
Li a spol. pripisujú tieto vplyvy na mozog zmenám v cytokinálnych úrovniach a imúnnej polarizácii (Th1/Th2 polarizácia), ktorá je podnietená vakcínami. Hliníkové adjuvanty spôsobujú Th2 polárizáciu. Li a spol. tvrdia, že ich výsledky naznačujú, že interakcia vakcín spôsobuje efekty aktivácie imunity:
„naša údaje naznačujú, že kombinácie rôznych vakcín môžu pôsobiť rôzne interakcie (zlepšovať alebo zhoršovať). Mechanizmus modulácie synaptickej plasticity prostredníctvom neonatálnej BCG/HBV vakcinácie môže spočívať v systemickej Th1/Th2 vychýlenosti doprevádzanej špecifickým profilom cytokínov a neurotropfínov v mozgu.“ (Li 2015)
Li 2015 preukázal, že vakcíny ovplyvňujú vývin mozgu mechanizmom aktivácie imunity. Ďalej, nakoľko hliníkové adjuvanty podnecujú Th2 aktiváciu a dlhodobú Th2 polarizáciu, výsledky Li 2015 naznačujú, že všetky vakcíny obsahujúce hliník ako adjuvant môžu spôsobiť negatívne efekty podobné HBV vakcíne. Napodobne výsledky Li 2015 načrtávajú, že štúdie preukázujúce, že aktivácia imunity je príčinou neurologických/psychiatrických porúch, sú relevantné ako nežiadúce účinky vakcín.
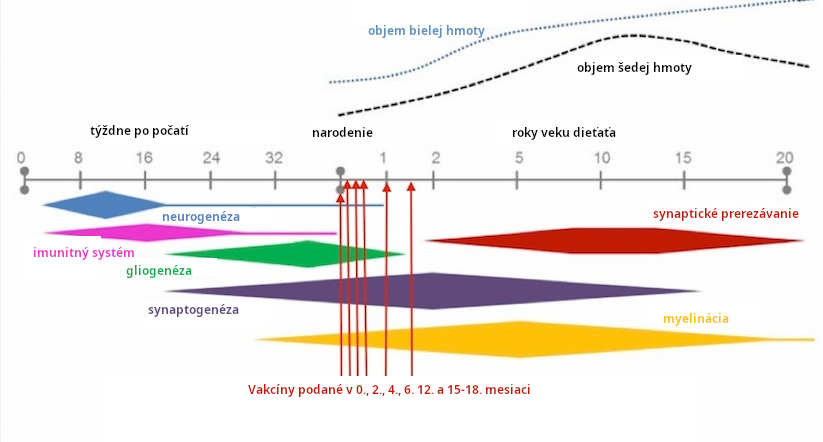
Vakcíny sú podávané počas synaptogenézy
Ďalší spôsob, ako zodpovedať otázku mozgovej zraniteľnosti voči aktivácii imunity je vziať do úvahy typy procesov mozgového vývinu, ktoré sa vyskytujú počas doby, keď sú podávané vakcíny. Vakcíny sú primárne podávané v prvých 18 mesiacoch po narodení. Ľudský mozog precháda intenzívnym a rapídnym vývinom počas tejto etapy. Synaptogenéza (formácia synaptických prepojení medzi neurónmi) je špeciálne intenzívna v tomto období.
Zraniteľnosť vyvíjajúceho sa mozgu voči aktivácii imunity zjavne súvisí s výskytom špecifických typov procesov mozgového vývinu (Tau 2010, Meyer 2006, Meyer 2007). Tieto procesy zahŕňajú migráciu (pohyb neurónov do finálneho miesta v mozgu), adhéziu (formáciu chemicko-mechanických väzieb medzi bunkami mozgu) a synaptogenézu (formáciu synaptických prepojení medzi neurónmi) medzi inými (neurogenéza, gliognéza, myelinácia atď.).
Cytokíny vplývajú na procesy vývinu mozgu. Napr. zvýšený IL-6 ovplyvňuje migráciu, adhéziu a synaptogenézu (Wei 2011). Zvýšený IL-6 v postnatálnom období vytvára exces excitačných synapsií a deficit inhibičných synapsií, a teda vedie k správaniam, typickým pre autizmus. (Wei 2012(a))
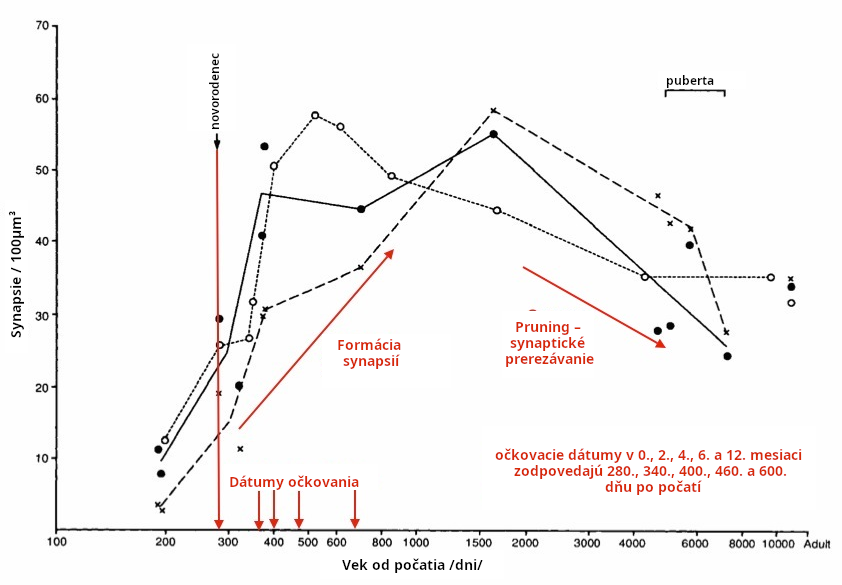
U ľudí, dramatické zvýšenie v synaptogenéze začína okolo času narodenia a pokračuje do veku 3 rokov (Huttenlocher 1997, Tau 2010, Stiles 2010, Semple 2013). Vakcíny sú podávané počas intenzívnej synaptogenézy. Pozri obr. 5-6. Zvýšený mozgový IL-6 podnietený očkovaním počas synaptogenézy môže spôsobiť excitačno-inhibičnú nerovnováhu vyklonenú smerom k excitácii. Excitačná nerovnováha je pozorovaná v ľudskom autizme (Robertson 2016, Freyberg 2015).
Synaptogenéza postupne spomaľuje počas detstva a dospievania. Tento fakt môže vysvetliť, prečo niektoré staršie deti a dospievajúci môžu trpieť autistickou regresiou po intenzívnom zápale mozgu, ale zjavne sa s vekom stávajú menej zraniteľnými na poškodenie aktiváciou imunity. Intenzívna synaptogenéza sa vyskytuje vo veku 0-18 mesiacov, keď je podávaných veľa vakcín. Následne vakcíny môžu mať negatívny dopad na synaptogenézu, ak podnietia zápal alebo IL-6 v mozgu.
Načasovanie procesov vývinu mozgu u ľudí podporuje domnienku, že ľudský mozog je zraniteľný voči aktivácii imunity a cytokínom v prvých rokoch života po narodení, keď sú podávané vakcíny. Narušenie synaptogenézy vakcínami podnietenou aktiváciou imunity je špecifickým problémom.
Hliníkové adjuvanty: Neurotoxicita v dávkach vakcín
Hliníkový (Al) adjuvant má podstatnú rolu v mnohých vakcínach: stimulovať aktiváciu imunity. Bez Al adjuvantov efektívnosť týchto vakcín by bola značne zredukovaná.
Hliníkové adjuvanty pozostávajú zo submikronických častíc (primárne častice) hliníkových zlúčenín, typicky AlOH, AlPO4, AlSO4 alebo zmiešanín. Primárne častice sú typicky združené do väčších častíc s veľkosťami okolo 2-20 mikrónov (Harris 2012). Hliníkové materiály ako adjuvant majú nízku rozpustnosť vo vode a v telesných tekutinách. Častice hliníkových adjuvantov sú biotrvanlivé a môžu zotrvať v tele mesiace alebo roky (Flarend 1997, Khan 2013, Cherardi 2001).
Hliník požitý v strave má nízku orálnu absorpciu (okolo 0,3%), je rýchlo vylúčený obličkami, je vo väčšine vylúčený z mozgu krvno-mozgovou bariérou a je v rozpustnej Al3+ ionickej forme (nie čiastočkovej). Tieto obrany sú adekvátne pre ochranu mozgu pred prirodzenými mierami požitia hliníka. Tieto ochranné mechanizmy sú neschopné chrániť mozog pri injekčnom podaní častíc hliníkového adjuvantu. Častice Al adjuvantu sú príliš veľké, aby boli odstránené obličkami a sú prenesené naprieč krvno-mozgovou bariérou makrofágmi.
Dávky hliníkových adjuvantov prijímané bábätkami podľa CDC očkovacieho kalendára sú:
Narodenie (Hep B vakcína prvých 24 hodín): 74 mcg/kg (250 mcg na 3,4 kilové dieťa)
2. mesiac: 245 mcg/kg (1225 mcg na 5 kilové dieťa)
4. mesiac: 150 mcg/kg (975 mcg na 6,5 kilové dieťa)
6. mesiac: 153 mcg/kg (1225 mcg na 8 kilové dieťa)
Toto sú maximálne možné dávky (pretože rôzne očkovacie produkty majú rôzne množstvá) pre deti s priemernou váhou.
Hromadenie dôkazov ukazuje, že hliníkové adjuvanty majú nežiadúce neurologické účinky pri dávke nižšej alebo približne rovnakej ako sú dávky, ktoré deti dostanú z vakcín. Tieto účinky sa zdá, že závisia od konkrétnej povahy a biotrvanlivosti hliníkového adjuvantu. Injekčný hliníkový adjuvant má nežiadúce účinky, ktoré sú očivine prenášané časticami a nezávislé od rozpustných Al3+ iónov uvoľnených pomalými rozložením častíc (Creapeaux 2017).
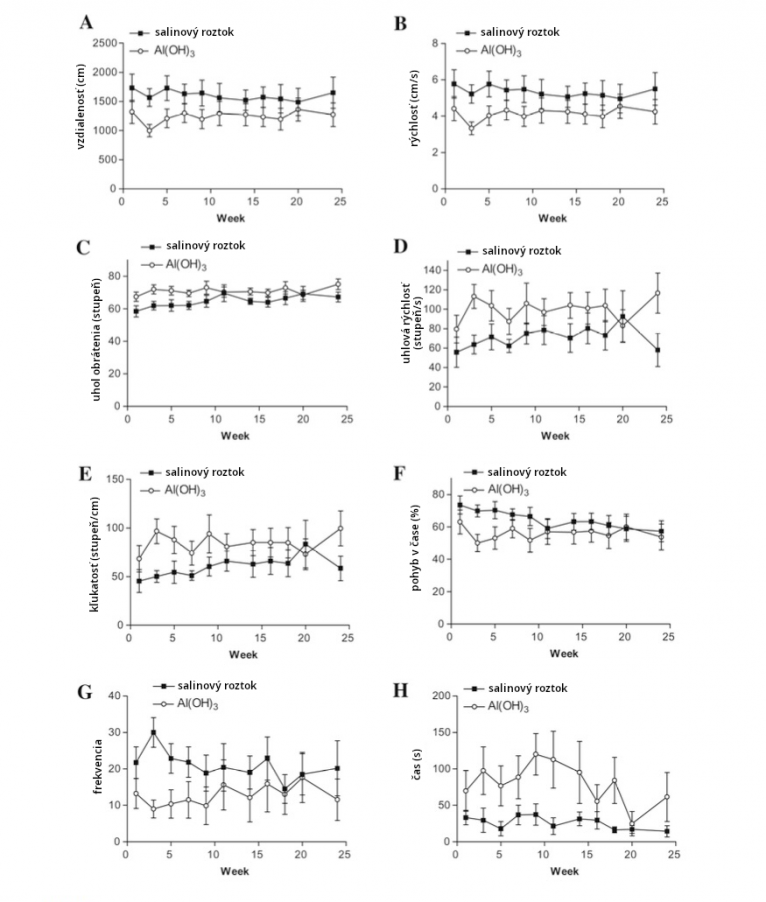
Injekcie s Al adjuvantami umyšíspôsobujú nežiadúce účinky pri dávkach ako sú relevantné pre vakcíny – 100, 200, 300 a 550 mcg/kg telesnej váhy (Crepeaux 2017, Shaw 2009, Petrik 2007, Shaw 2013). Tie zahŕňajú deficity v učení a pamäti (Shaw 2009), deficity v neuromuskulárnej sile(funkcii (Petrik 2007) a zmeny v lokomotorickej aktivite a alebo v držaní tela (Shaw 2009, Shaw 2013). Autizmus sa spája s abnormalitami v držaní tela a pohybe (Kindregan 2015) a s pamäťovou disfunkciou (Williams 2006).
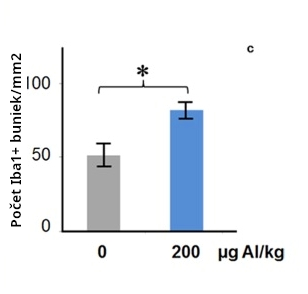
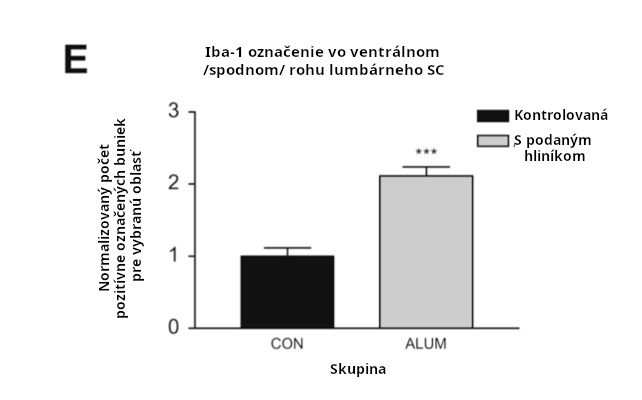
Dávky 200mcg/kg ako 3x66mcg/kg (Creapeaux 2017) a 300mcg/kg ako 6x 50mcg/kg (Shaw 2009) hliníkového adjuvantu zvýšili aktiváciu mikroglií v prednej časti predného mozgu a zároveň v lumbárnej mieche. Zvýšená aktivácia mikroglií bola nameraná asi 6 mesiacov po podaní injekcie s Al adjuvantom, čo naznačuje, že aktivácia mikroglií je chronická. Aktivované mikroglie naznačujú postupujúci zápalový proces a prítomnosť zvýšených cytokínov. Ľudský autizmus má aktivované mikrogliá a zvýšené cytokíny naprieč mozgom (Vargas 2005, Suzuki 2013, Li 2009).
Aktivované mikrogliá sú implikované ako kauzálny faktor v autizme, pretože mikrogliá sprostredkuvávajú zápal v mozgu. Mikrogliá môžu produkovať IL-6, keď sú v aktivovanom stave. Nedávna recenzia o mikrogliách a autizme (Takano 2015) tvrdí:
„akékoľvek faktory, ktoré menia počet alebo aktivačný stav mikroglií buď v maternici alebo počas skorého postnatálneho obdobia môžu značne ovplyvniť neurálny vývin, a tak vyústiť do neurovývinových porúch, vrátane autizmu.“ (Takano 2015)
Zdá sa, že mikrogliá hrajú dôležitú úlohu v súvislosti s autizmom (Takano 2015, Kneusel 2014). Preto aktivácia mikroglií, zapríčinená hliníkovými adjuvantami, naznačuje rolu v autizme.
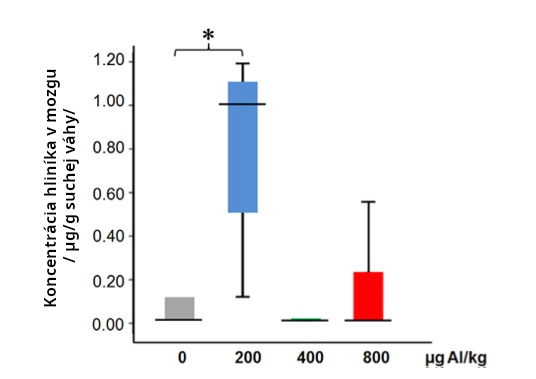
Niekoľko štúdií ukázalo, že Al adjuvanty zvyšujú obsah hliníka v mozgu (Crepeaux 2017, Flarend 1997, Shaw 2009, Khan 2013, Crepeaux 2015). Dávka 200mcg/kg hliníkového adjuvantu spôsobila 50-násobný nárast v obsahu hliníka v mozgu myší, od 0,02 ug/g do 1.0 ug/g suchej váhy mozgu (Crepeaux 2017). Tieto merania boli vykonané 6 mesiacov po poslednej injekcii naznačujúc, že Al pretrváva v mozgu dlhodobo (Crepeaux 2017). Pozri obr. 10. Bolo zistené, že Al adjuvanty sa hromadia v mozgu myší do jedného roka po injekcii (Khan 2013). Crepeaux 2015 ukazuje trvácnosť a zvyšujúce sa hromadenie častíc Al adjuvantov do 270 dní v slezine a v lympatických uzlinách myší. Zvýšené hromadenie Al vo vzdialených orgánoch po čase naznačuje, že toxické efekty sa možu časom zvyšovať a tak byť oneskorené o mesiace alebo roky po podaní.
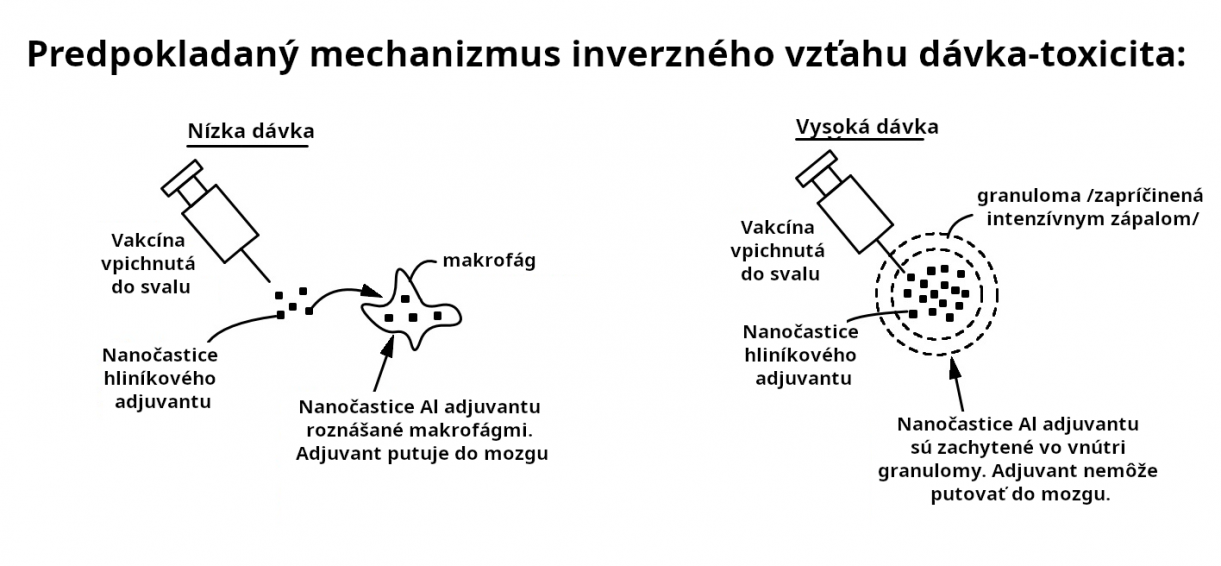
Dávky 400 a 800mcg/kg v štúdii Crepeaux 2017 nespôsobovali nežiadúce účinky alebo zvýšený mozgový hliník. Tento prekvapivo zvrátený vzťah dávky a reakcie pripisujú autori granulomám indukovaným pri vyšších dávkach. Granulomy zachytia Al adjuvant akoby do pasce na mieste injekčného podania, aby tak predišli transportu do mozgu a ďalších citlivých tkanív. Granulomy sa vyskytujú po zhruba 1% očkovaní (Bergfors, 2014). To je dôvodom pre obavu, keďže to naznačuje, že u 99% očkovaní hliníkový adjuvant môže byť transportovaný po celom tele. Nie je uzavretý v granulome. Pozri obr. 11.
Transport častíc a markofágový chemotaktický proteín MCP-1
Hliníkové adjuvanty cestujú do mozgu (Khan 2013, Creapeaux 2015, Crepeaux 2017, Shaw 2009, Flarend 1997). Častice Al adjuvantu sú prenášané cez krvno-mozgovú bariéru a do mozgu makrofágmi (Khan 2013). Transport zabezpečuje makrofágový chemotaktický proteín MCP-1 (Khan, 2013). MCP-1 spôsobuje, že makrofágy cestujú po tele a do mozgu. Transport častíc do mozgu makrofágmi je dobre potvrdený a bol skúmaný pre terapeutické využitia (Choi 2012, Pang 2016).
MCP-1 je zvýšený v mozgoch ľudských autistov (Vargas 2005) a je zvýšený v krvi novorodencov neskôr diagnostikovaných na autizmus (Zerbo 2014). To naznačuje, že novorodenci s vysokým MCP-1 budú mať ešte vyššiu mieru transportu Al adjuvantu do mozgu, keď im budú vpichnuté Al adjuvanty vo vakcínach. Toto je konzistentné s tým, žeAl adjuvant spôsobujú autizmus podnietením aktivácie imunity a zvýšením cytokínov v mozgu.
Hliník podnecuje expresiu IL-6 v mozgu
Vo voderozpustné hliníkové soli napr. AlCl3, Al laktát, podnecujú zvýšený IL-6 v mozgu a ďalších tkanivách. Fakticky sa zdá, že hliník selektívne podnecuje IL-6 (Viezeliene, 2013). Štúdie o pôsobení hliníka a IL-6 expresie v mozgu zahŕňajú:
Cao 2016: Požitie 30 alebo 90mg/kg/deň hliníka (vo forme AlCl3) počas 90 dní viedlo k významnému zvýšeniu genovej expresie IL-6 a ďalších cytokínov v mozgu (hippokampus).
Alawdi 2016: Požitie 3,4mg/kg/deň hliníka vo forme AlCl3 počas 6 týždňov viedlo k 4-násobnému zvýšeniu IL-6 v mozgu (hippokampus). Táto dávka je oveľa nižšia ako zastaralý limit, tzv. „úroveň žiadnych pozorovaných nežiadúcich účinkov“ NOAEL pre ústne dávky (26 a 62mg/kg/deň), používaná ako hranica toxicity (Mitkus 2011, Offit 2003).
V podstate ďalšie experimenty ukazujú, že ústne dávky 3.4, 4, 5.6, 6 a 20.2 mg/kg/deň hliníka spôsobujú početné nežiadúce účinky u myší alebo potkanov a napriek tomu NOAEL pre ústne požitý hliník je v súčasnosti neznámy (Alawdi 2016, Dera 2016, Sethi 2008, Bilkei-Gorzo 1993). Môže sa spustiť Il-6 preto, lebo hliník silno spúšťa oxidatívny stres (Exley 2003). Oxidatívny stres spúšťa expresiu IL-6 (Viezeliene 2013).
CDC webstránka cituje fatálne chybné štúdie o hliníkových adjuvantoch (Mitkus 2011)
Dávky Al adjuvantov prijímané bábätkami sa dramaticky zvýšili, keď sa rozšíril očkovačí kalendár v 80-tych a 90-tych rokoch. Avšak, ako sa rozšíril očkovací kalendár, zvýšené dávky Al adjuvantov neboli testované ohľadom ich bezpečnosti. Vládne agentúry (HHS, NIH, CDC, FDA) nezaujímali žiadne nové experimentálne projekty o toxicite Al adjuvantov.
Na podporu bezpečnosti Al adjuvantov pri dnešných vysokých dávkach, CDC cituje štúdiu FDA z roku 2011 ohľadom vplyvu hliníka vo vakcínach (Mitkus 2011). Táto štúdia je jediným vedeckým dôkazom, o ktorý sa v citátoch na webstránkach opiera CDC a FDA, na podporu bezpečnosti Al adjuvantov.
Mitkus 2011 je štúdia teoretického modelu kinetiky Al adjuvantu, neobsahuje žiadne nové údaje ohľadom toxicity Al adjuvantov (zo zvieracich modelov alebo epidemiológie). Mitkus 2011 prepočítava telesnú záťaž hliníka ako sa postupne uvoľňujú Al častice a porovnáva záťaž tela rozpusteným hliníkom k „minimálnej rizikovej miere“ - MRL. MRL je odvodené zo štúdie o toxicite požitého hliníka u myší (Golub 2001). Golub 2001 udáva NOAEL 26mg/kg/deň pri požití, čo je prepočítané na MRL u detí v miere 1mg/kg/deň pri použití bezpečnostného faktoru okolo 30.
Mitkusova štúdia je fatálne chybná pre tieto dôvody:
1. Mitkus sa domnieva, že všetky častice adjuvantov sú neškodné
Mitkus zakladá svoju štúdiu na domnienke, hoci ju netvrdí, že Al adjuvanty majú nulovú toxicitu vo forme častíc. Mitkus pripúšťa len potenciálnu toxicitu hliníkových iónov Al3+ uvoľnených pomalým rozpúšťaním častíc Al adjuvantu.
Al adjuvanty pozostávajú z nízko rozpustiteľných a biologicky trvácich mikroskopických častíc. Mitkusova analýza predpokladá, že častice sú absolútne netoxické a dokonale neškodné, dokonca aj keď sú prítomné v mozgu a ďalších orgánoch. Mitkus neposkytuje žiadne zdôvodnenie pre túto netvrdenú domnienku. Ďalej táto domnienka protirečí nedávnym zisteniam o toxicite Al adjuvantov (Crepeaux 2017) a toxicite častíc vo všeobecnosti. Častice môžu mať toxický vplyv spôsobený povrchovou chémiou (napr. povrchový náboj a povrchová katalytická aktivita) a tvarom častíc, okrem iného charakteristikami pevných častíc (Sharifi 2012, Podila 2013).
Niekoľko štúdií ukazuje, že injekčne podané Al adjuvanty spôsobujú abnormality správania, abnornálne prírastky váhy, narušenie učenia a pamäte, apoptózu – úhyn motorických neurónov, deficity neuromuskulárnej sily, chronickú mikrogliálnu aktiváciu, mozgový zápal a ohromný (napr. 50-násobný) nárast obsahu hliníka v mozgu a mieche (Petrik 2007, Shaw 2009, Shaw 2013, Crepeaux 2017). Tieto nežiadúce účinky sa vyskytujú pri dávkach menej alebo približne rovnako ako je dávka prijímaná bábätkami podľa CDC očkovacieho kalendára.
2. Nový výskum uvádza požité dávky hliníka nižšie ako 26mg/kg/deň ako škodlivé
Mitkus predpokladá, že toxicita Al adjuvantov je možná výlučne kvôli rozpustiteľnému hliníku (Al3+ iónom(, uvoľnenému postupným rozpúšťaním častíc Al adjuvantu. Pre určenie hranice miery toxicity rozpustného Al sa Mitkus opiera o štúdiu, v ktorej boli kŕmené myši (Golub 2001) a žiadne pozorované nežiadúce účinky NOAEL pri ústnych dávkach 26mg/kg/deň požitého hliníka neboli hlásené. Mitkus použil bezpečnostný faktor 30x pre aplikáciu tejto dávky pre ľudí, čo sa zdá byť rozumné.
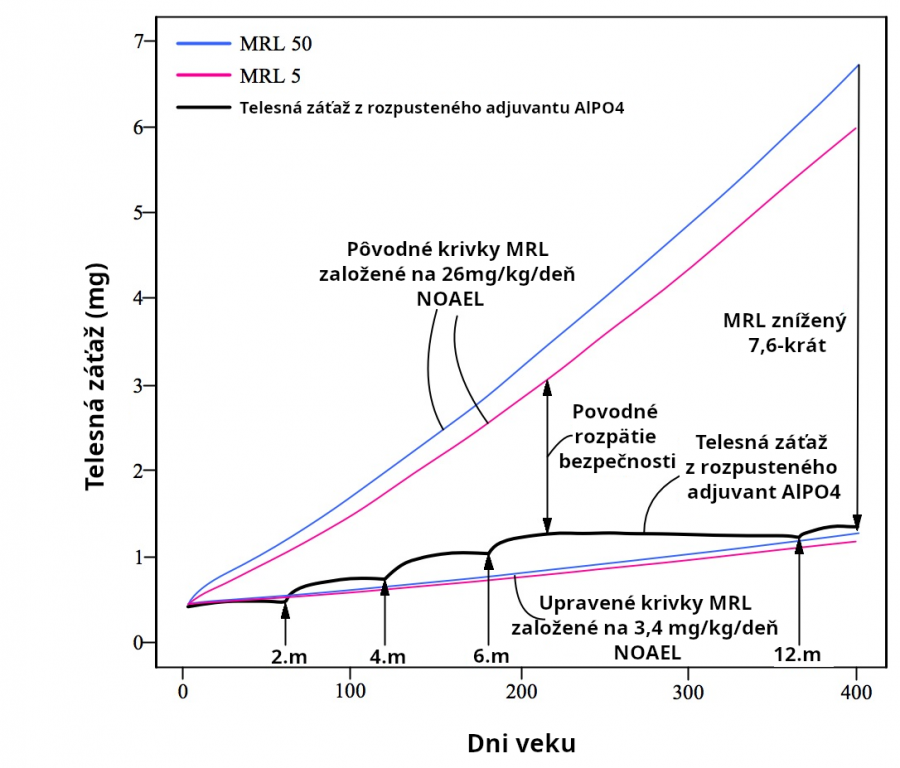
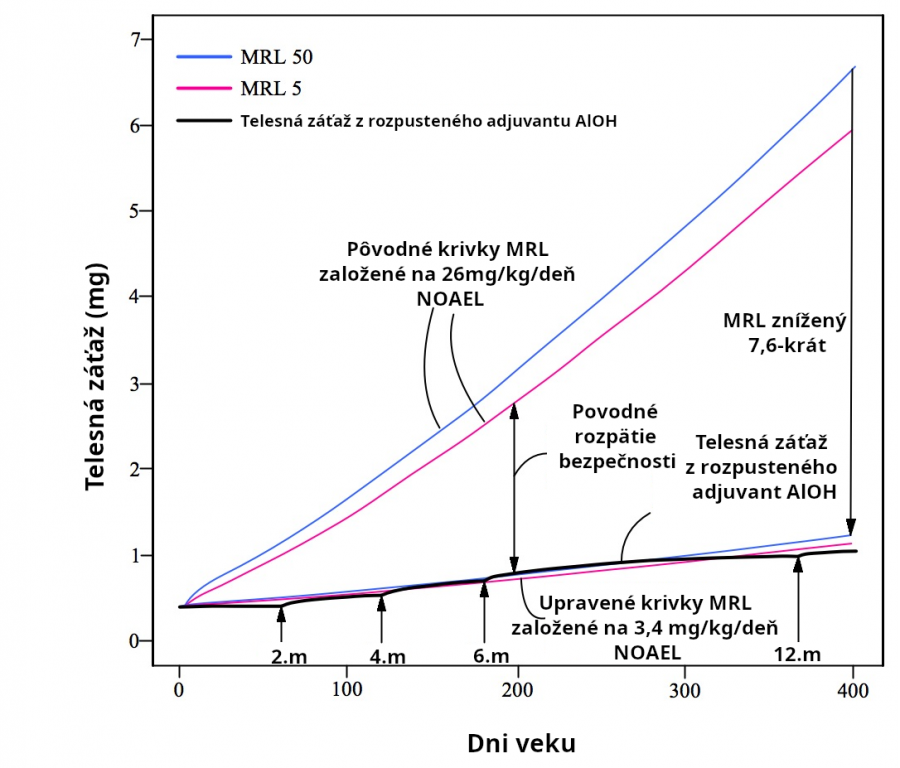
Avšak ďalšie experimenty ukazujú, že oveľa menšie orálne dávky 3.4, 4, 5.6, 6 a 20.2mg/kg/deň hliníka môžu spôsobiť nežiadúce účinky u myší a potkanov (Alawdi 2016, Dera 2016, Sethi 2008, Sethi 2009, Bilkei-Gorzo 1993). Nežiadúce účinky zahŕňajú chronický zápal mozgu, poškodenie učenia a pamäte a zápal obličiek. Takže Mitkusova analýza je nesprávna, lebo 26mg/kg/deň nie je NOAEL. Minimálna riziková miera MRL stanovená Mitkusom je príliš vysoká o faktor najmenej 26/3.4 = 7.6. Keď použijeme upravený NOEAL 3,4 mg/kg(deň (na základe Alawdi 2016), výsledkom je, že hliník vo vakcíne presahuje MRL pre AlPO4 adjuvant a približne je rovnaký ako MRL pre AlOH adjuvant. Nové upravené MRL miery naznačujú, že adjuvant hliníkového fosfátu (Obr. 12) a adjuvant hliníkového hydroxidu (Obr. 13) z CDC očkovacieho kalendára môžu spôsobiť toxicitu rozpustiteľným hliníkom per se.
Nakoľko 3,4 mg/kg/deň nie je NOAEL (nežiadúce účinky boli pozorované pri tejto dávke), skutočný NOAEL je menej ako 3,4mg/kg/deň. Pozri obr. 12-13.
3. Žiadne údaje o toxicite Al adjuvantov nie sú citované, hoci boli dostupné
Mitkus necituje žiadne údaje o toxicite injekčne podávaných Al adjuvantov. Namiesto toho používa dáta o toxicite požitého, nečasticového, vo vode rozpustiteľného hliníka (štúdia Golub 2001 použila Al laktát) na určenie MRL. Jeho údaj má pôvod v jedinej štúdii (Golub 2001).
Takže je pozorohudné, že Mitkus deklaruje bezpečnú hladinu injekčne podaného Al adjuvantu bez citovania údajov o toxicite Al adjuvantu. Chyba nie je nevyhnutná a zanedbateľná, pretože najmenej dve štúdie o toxicite injekčne podaného Al adjuvantu u zvierat existovali pred publikáciou Mitkesovej štúdie v roku 2011 (Petrik 2007, Shaw 2009). Tieto štúdie neboli citované alebo zmienené v štúdii Mitkes 2011.
Každá z týchto troch chýb je fatálna pre právoplatnosť Mitkesovej štúdie o prehlásení hliníkových adjuvantov za bezpečné. CDC úplne chýba akýkoľvek dôkaz o bezpečnosti Al adjuvantov. To je špeciálne pravda o bezpečnosti ohľadom neurologických a dlhodobých účinkov, pretože ďalšie dostupné štúdie o bezpečnosti Al adjuvantov (napr. Jefferson 2004) neberú ohľad na (alebo nie sú schopné rozoznať) tieto účinky.
CDC zlyhalo v preskúmaní toxicity hliníkových adjuvantov
CDC nevykonalo žiadne epidemiologické štúdie o dlhodobej bezpečnosti (napr. zvažujúc neurologické účinky) hliníkových adjuvantov. Existuje jedna ekologická štúdia údajov na národnej úrovni, ktorá potvrdila súvislosť Al adjuvantov a autizmu (Tomlienovic 2011). Tým, že ide o ekologickú štúdiu, je vysoko náchylná k predpojatosti.
Dr. Frank DeStefano z CDC úradu pre bezpečnosť vakcín je spoluautorom projektovej štúdie (Glanz 2015) o používaní systému Vaccine Safety Datalink VSD na preskúmanie bezpečnosti jednotlivých zložiek vakcín. Táto štúdia sa zameriava na Al adjuvanty. Uznáva, žethimerosoal je jediná zložka vakcín študovaná s ohľadom na autizmus alebo neurologickú bezpečnosť a že možná súvislosť Al adjuvantov a autizmu nebola skúmaná v epidemiologických štúdiách. Glanz 2015 tvrdí: „Dodnes neexistujú populačné štúdie špecificky dizajnované na vyhodnotenie súvislosti klinicky zmysluplných účinkov a neantigénych zložiek, okrem thimerosalu.“
CDC neprešetrovalo obavy o bezpečnosť hliníkových ajduvantov napriek hromadiacim sa vedeckým dôkazom o škodlivosti a dôkazom súvislosti Al adjuvantov a mechanizmov imúnnej aktivácie mozgových poškodení.*
Záver
Tu zhrnutá veda nám hovorí konzistený a silný príbeh, že vakcíny môžu spôsobiť autizmus stimuláciou aktivácie imunity a zvýšených cytokinov v mozgu. Al ajuvanty sú považované za príčinu autizmu, pretože môžu putovať do mozgu, spôsobujú aktiváciu mikroglií v dávkach ako sú vo vakcínach a preto, lebo hliník podnecuje IL-6 v mozgu.
V tvrdeniach, že neexistuje súvislosť vakcín a autizmu, CDC cituje vedecký dôkaz, ktorý nie je relevantný pre bezpečnosť Al adjuvantov alebo nie je schopný vyvrátiť súvislosť medzi Al adjuvantami a autizmom (Taylor 2014, De Stefano 2013,, Mitkus 2011). Na podporu svojho tvrdenia sa CDC opiera o značne chybnú štúdiu teoretického modelovania (Mitkus 2011). Existujú nepatrné vedecké dôkazy na podporu bezpečnosti Al adjuvantov, hlavne čo sa týka autizmu a ďalších dlhodobých neurologických účinkov.
*Avšak Glanzova štúdia poznamenáva, že skúmanie hliníkových adjuvantov je problematické kvôli očakávaným malým rozdielom v príjme v skupinách s nízkych a vysokým príjmom. Glanz 2015 zhrňuje: „deti pod 10. percentilo by prijímali medzi 0 a 3,1mg, kým deti nad 90. percentilom by prijímali medzi 4,8 a 5,3 mg hliníka vo vakcínach. Nie je jasné, či takéto rozdiely v príjme hliníka by boli biologicky významné“ (Glanz 2015). Takže epidemiologické štúdie nemôžu poskytnúť spoľahlivý dôkaz o bezpečnosti alebo škodlivosti. Kontrolované, výhľadové ľudské testy o príjme hliníkových adjuvantov z vakcín budú pravdepodobne zakázané pre etické dôvody. Taktiež Al adjuvanty sú podstatné zložky u vakcín s Al adjuvantom. Následne by bolo veľkou výzvou nadizajnovať štúdie dlhodobých nežiadúcich účinkov Al adjuvantov u ľudí. Experimenty na zvieratách môžu poskytnúť hodnotné informácie. Al adjuvanty by mali byť testované pre účinky na: 1. excitačnú/inhibičnú nerovnováhu, 2. základné symptómy autizmu (sociálne, komunikačné a repetatívno-stereotypné správania), 3. IL-6, IL-17 a hladiny ďalších cytokínov v mozgu, 4. ďalšie fyziologické abnormality súvisiace s autizmom (napr. mitochondriálna dysfunkcia, dysbióza mikrobiómu, strata Purkiněho buniek, abnormality mozočka, atď.) a 5. aktiváciu mikroglií a imúnnu aktivitu v mozgu. Preskúmanie týchto účinkov môže poskytnúť hodnotné informácie týkajúce sa bezpečnosti Al adjuvantov.
Zdroje
Akintunde et al., 2015 Increased production of IL-17 in children with autism spectrum disorders and comorbid asthma, Journal of Neuroimmunology 286 (2015) 33-41.
Al-Ayadhi et al., 2012 Elevated serum levels of interleukin-17A in children with autism, Journal of Neuroinflammation 2012, 9:158.
Alawdi et al., Neuroprotective Effect of Nanodiamond in Alzheimer’s Disease Rat Model: a Pivotal Role for Modulating NF-κB and STAT3 Signaling, Molecular Neurobiology, 54 (3):1906-1918.
Atladottir et al., Maternal infection requiring hospitalization during pregnancy and autism spectrum disorders, Journal of Autism and Developmental Disorders, 2010 Dec;40(12):1423-1430.
Bauman et al., 2014 Activation of the Maternal Immune System During Pregnancy Alters Behavioral Development of Rhesus Monkey Offspring, Biological Psychiatry, 2014;75: 332–341
Bergfors et al., 2014 How common are long-lasting, intensely itching vaccination granulomas and contact allergy to aluminium induced by currently used pediatric vaccines? A prospective cohort study, European Journal of Pediatrics, 173:1297–1307.
Bilkei-Gorzo, 1993, Neurotoxic effect of enteral aluminum, Food and Chemical Toxicology, 31(5):357-361.
Brown et al., 2014 Metabolic consequences of interleukin-6 challenge in developing neurons and astroglia, Journal of Neuroinflammation, 11:183.
Brown et al., Epidemiologic studies of exposure to prenatal infection and risk of schizophrenia and autism, Developmental Neurobiology, 2012 October ; 72(10): 1272–1276.
Bruce et al., 2011 Converging pathways lead to overproduction of IL-17 in the absence of vitamin D signaling, 2011 Aug; 23(8): 519–528.
Careaga et al 2017 Maternal Immune Activation and Autism Spectrum Disorder: From Rodents to Nonhuman and Human Primates, Biological Psychiatry, March 1, 2017; 81:391–401.
Chen et al., Postnatal systemic inflammation exacerbates impairment of hippocampal synaptic plasticity in an animal seizure model, Neuroimmunomodulation, 2013;20(4):223-32.
Choi et al., 2012, Delivery of nanoparticles to brain metastases of breast cancer using a cellular Trojan horse, Cancer Nanotechnology, 3:47–54.
Choi et al., 2016 The maternal interleukin-17a pathway in mice promotes autismlike phenotypes in offspring, Science, 2016 Feb 26; 351(6276): 933–939.
Ciaranello et a The Neurobiology of Infantile Autism, The Neuroscientist, 1:361-367
Coiro et al., Impaired synaptic development in a maternal immune activation mouse model of neurodevelopmental disorders, Brain, Behavior, and Immunity, Nov;50:249-258.
Crepeaux et al., 2015 Highly delayed systemic translocation of aluminum-based adjuvant in CD1 mice following intramuscular injections, Journal of Inorganic Biochemistry, 152:199-205.
Crepeaux et al., 2017 Non-linear dose-response of aluminium hydroxide adjuvant particles: Selective low dose neurotoxicity, Toxicology, 375 (2017) 48–57.
DeLong et al., 1981 Acquired reversible autistic syndrome in acute encephalopathic illness in children, Archives of Neurology, 36:191-194.
Dera 2016, Protective effect of resveratrol against aluminum chloride induced nephrotoxicity in rats, Saudi Medical Journal, 37 (4).
DeStefano et al., 2013 Increasing Exposure to Antibody-Stimulating Proteins and Polysaccharides in Vaccines Is Not Associated with Risk of Autism, The Journal of Pediatrics, 163 (2).
Deverman and Patterson, 2009 Cytokines and CNS Development, Neuron 64:61-78.
Drozdenko et al., 2014 Oral vitamin D increases the frequencies of CD38+ human B cells and ameliorates IL-17-producing T cells, Experimental Dermatology, 23: 107-112.
Estes and McAllister, 2016 Maternal immune activation: implications for neuropsychiatric disorders, Science, 353 (6301) 772-777.
Exley, 2003 The Pro-Oxidant Activity of Aluminum, Free Radical Biology and Medicine, 36(3): 380-387.
Flarend et al., 1997 In vivo absorption of aluminum-containing vaccine adjuvants using 26 Al, Vaccine, 15(12/13):1314-1318.
Freyberg et al., 2015 Reduced perceptual exclusivity during object and grating rivalry in autism, Journal of Vision, 15(13):11, 1–12.
Galic et al., 2008 Postnatal Inflammation Increases Seizure Susceptibility in Adult Rats, The Journal of Neuroscience, 2008, 28 (27) 6904-6913.
Garay et al., 2013 Maternal immune activation causes age- and region-specific changes in brain cytokines in offspring throughout development, Brain, Behavior, and Immunity, 31: 54-68.
Ghaziuddin et al., 2002 Autistic symptoms following herpes encephalitis, European Child and Adolescent Psychiatry, Vol. 11, No. 3:142-146.
Gherardi et al., 2001 Macrophagic myofasciitis lesions assess long-term persistence of vaccine-derived aluminium hydroxide in muscle, Brain, 124:1821-1831.
Ghiani et al., 2011 Early effects of lipopolysaccharide induced inflammation on foetal brain development in rat, ASN Neuro, 3 (4): 233-245.
Gillberg 1986 Brief Report: Onset at Age 14 of a Typical Autistic Syndrome. A Case Report of a Girl with Herpes Simplex Encephalitis, Journal of Autism and Developmental Disorders, VoL 16, No. 3:369-375.
Giulivi et al 2013 Gestational Exposure to a Viral Mimetic Poly(I:C) Results in Long- Lasting Changes in Mitochondrial Function by Leucocytes in the Adult Offspring, Mediators of Inflammation, Vol 2013:609602.
Glanz et al., 2015, Cumulative and episodic vaccine aluminum exposure in a population- based cohort of young children, Vaccine 33:6736–6744.
Golub et al., 2001 Long-term consequences of developmental exposure to aluminum in a suboptimal diet for growth and behavior of Swiss Webster mice, Neurotoxicology and Teratology 23 (2001) 365–372.
Gupta et al., 1998 Th1- and Th2-like cytokines in CD4+ and CD8+ T cells in autism, Journal of Neuroimmunology, 85:106-109.
Harre et al., 2008 Neonatal inflammation produces selective behavioural deficits and alters N-methyl-D-aspartate receptor subunit mRNA in the adult rat brain, European Journal of Neuroscience, 2008 Feb; 27(3): 644–653.
Harris et al., 2012 Alhydrogel® adjuvant, ultrasonic dispersion and protein binding: A TEM and analytical study, Micron, 43:192-200.
Hsiao et al., 2013 The microbiota modulates gut physiology and behavioral abnormalities associated with autism, Cell, 155(7): 1451-1463.
Huttenlocher and Dabholkar, 1997 Regional Differences in Synaptogenesis in Human Cerebral Cortex, Journal of Camparative Neurology, 387:167–178 (1997).
Jefferson 2004 Adverse events after immunisation with aluminium-containing DTP vaccines: systematic review of the evidence, The Lancet 4:84-90.
Jones et al., 2016 Autism with Intellectual Disability is Associated with Increased Levels of Maternal Cytokines and Chemokines During Gestation, Molecular Psychiatry, 22(2):273-279.
Khan et al., 2013 Slow CCL2-dependent translocation of biopersistent particles from muscle to brain, BMC Medicine, 11:99.
Kindregan et al., 2015 Gait Deviations in Children with Autism Spectrum Disorders: A Review, Autism Research and Treatment, ID:741480.
Knuesel et al., 2014, Maternal immune activation and abnormal brain development across CNS disorders, Nature Reviews 10:643-660.
Labouesse et al., 2015, Long-term pathological consequences of prenatal infection: beyond brain disorders, American Journal of Physiology, 309:1.
Li et al. 2009 Elevated Immune Response in the Brain of Autistic Patients, Journal of Neuroimmunology, 207(1-2): 111–116.
Li et al., 2015 Neonatal vaccination with bacillus Calmette–Guérin and hepatitis B vaccines modulates hippocampal synaptic plasticity in rats, Journal of Neuroimmunology, 288 (2015) 1-12.
Machado et al., 2015 Maternal Immune Activation in Nonhuman Primates Alters Social Attention in Juvenile Offspring, Biological Psychiatry, 2015 May 1;77(9):823-32.
Malkova et al., 2012 Maternal immune activation yields offspring displaying mouse versions of the three core symptoms of autism, Brain Behaviorand Immunity, 2012 May ; 26(4): 607–616.
Marques et al., 2014 Autism Spectrum Disorder Secondary to Enterovirus Encephalitis, Journal of Child Neurology, 2014, Vol. 29(5) 708-714.
Mestas et al., 2004 Of Mice and Not Men: Differences between Mouse and Human Immunology, Journal of Immunology, 0022-1767:2731-2738.
Meyer et al., 2006 The Time of Prenatal Immune Challenge Determines the Specificity of Inflammation-Mediated Brain and Behavioral Pathology, The Journal of Neuroscience, 26(18):4752– 4762.
Meyer et al., 2007 The neurodevelopmental impact of prenatal infections at different times of pregnancy: the earlier the worse?, Neuroscientist, Jun;13(3):241-56.
Meyer et al., 2009 In-vivo rodent models for the experimental investigation of prenatal immune activation effects in neurodevelopmental brain disorders, Neuroscience and Biobehavioral Reviews, 33 (2009) 1061–1079.
Meyer 2014, Prenatal Poly(I:C) Exposure and Other Developmental Immune Activation Models in Rodent Systems, Biological Psychiatry, 75:307-315.
Mitkus et al., 2011 Updated aluminum pharmacokinetics following infant exposures through diet and vaccination, Vaccine 29 (2011) 9538–9543.
Offit et al., 2003 Addressing Parents’ Concerns: Do Vaccines Contain Harmful Preservatives, Adjuvants, Additives, or Residuals? Pediatrics, 112(6): 1394-1401.
Oskvig et al., 2012 Maternal immune activation by LPS selectively alters specific gene expression profiles of interneuron migration and oxidative stress in the fetus without triggering a fetal immune response, Brain Behavior and Immunity, 2012 May ; 26(4): 623–634.
Pang et al., 2016 Exploiting macrophages as targeted carrier to guide nanoparticles into glioma, Oncotarget 7(24):37081.
Parker-Athill and Tan, 2010 Maternal Immune Activation and Autism Spectrum Disorder: Interleukin-6 Signaling as a Key Mechanistic Pathway, NeuroSignals, 2010;18:113–128.
Petrik et al., 2007 Aluminum Adjuvant Linked to Gulf War Illness Induces Motor Neuron Death in Mice, NeuroMolecular Medicine, Vol. 9, 83-100.
Pineda et al., 2013 Maternal immune activation promotes hippocampal kindling epileptogenesis in mice, Annals of Neurology, 2013 July ; 74(1): 11–19.
Podila et al., 2013 Toxicity of Engineered Nanomaterials: A Physicochemical Perspective, Journal of Biochemical and Molecular Toxicology, 2013 January ; 27(1): 50–55.
Robertson et al., 2016 Reduced GABAergic Action in the Autistic Brain, Current Biology, 26, 1-6.
Saad et al., 2016 Vitamin D status in autism spectrum disorders and the efficacy of vitamin D supplementation in autistic children, Nutritional Neuroscience, 19 (8) 346-351.
Semple et al., 2013 Brain development in rodents and humans: Identifying benchmarks of maturation and vulnerability to injury across species, Progress in Neurobiology, Jul-Aug;106-107:1-16.
Sethi et al., 2008 Aluminium-induced electrophysiological, biochemical and cognitive modifications in the hippocampus of aging rats, Neurotoxicology 29, 1069-1079.
Sethi et al., 2009 Curcumin attenuates aluminium-induced functional neurotoxicity in rats, Pharmacology, Biochemisatry, and Behavior 93:31-39.
Shen et al., 2016 Postnatal activation of TLR4 in astrocytes promotes excitatory synaptogenesis in hippocampal neurons, Journal of Cell Biology, 215(5):719-734.
Sharifi et al., 2012 Toxicity of Nanomaterials, Chemical Society Reviews, 2012 Mar 21; 41(6): 2323–2343.
Shaw and Petrik, 2009 Aluminum hydroxide injections lead to motor deficits and motor neuron degeneration, Journal of Inorganic Biochemistry 103 (11).
Shaw and Tomljenovic, 2013 Administration of aluminium to neonatal mice in vaccine-relevant amounts is associated with adverse long term neurological outcomes, Journal of Inorganic Biochemistry, 128 (2013) 237–244.
Shi et al., 2009 Activation of the Maternal Immune System Alters Cerebellar Development in the Offspring, Brain, Behavior, and Immunity, January, 23(1): 116–123.
Smith et al., 2007 Maternal Immune Activation Alters Fetal Brain Development through Interleukin-6, Journal of Neuroscience, 2007 October 3; 27(40).
Smith et al., 2012, Maternal Immune Activation Increases Neonatal Mouse Cortex Thickness and Cell Density, Journal of Neuroimmune Pharmacology, 7(3):529-532.
Stiles et al., 2010 The Basics of Brain Development, Neuropsychology Reviews (2010) 20:327–348.
Suzuki et al., 2011 Plasma Cytokine Profiles in Subjects with High-Functioning Autism Spectrum Disorders, PloS ONE 6(5).
Suzuki et al., 2013 Microglial Activation in Young Adults With Autism Spectrum Disorder, JAMA Psychiatry 70(1): 49-58.
Informed Consent Action NetworkAutism & Aluminum Adjuvants in Vaccines Takano 2015 Role of Microglia in Autism: Recent Advances, Developmental Neuroscience, 37:195-202.
Tau and Peterson, 2010 Normal Development of Brain Circuits, Neuropsychopharmacology, (2010) 35:147–168.
Taylor et al., 2014 Vaccines are not associated with autism: An evidence-based meta-analysis of case-control and cohort studies, Vaccine, 32:3623-3629.
Tomljenovic and Shaw, 2011 Do aluminum vaccine adjuvants contribute to the rising prevalence of autism? Journal of Inorganic Biochemistry 105.
Tsilioni et al., 2015 Children with autism spectrum disorders, who improved with aluteolin-containing dietary formulation, show reduced serum levels of TNF and IL-6, Translational Psychiatry, 5, 647.
Vargas et al., 2005 Neuroglial Activation and Neuroinflammation in the Brain of Patients with Autism, Annals of Neurology, 2005;57:67–81.
Viezeliene et al., 2013 Selective induction of IL-6 by aluminum-induced oxidative stress can be prevented by selenium, Journal of Trace Elements in Medicine and Biology, 27:226-229.
Wei et al., 2011 IL-6 is increased in the cerebellum of autistic brain and alters neural cell adhesion, migration and synaptic formation, Journal of Neuroinflammation 2011, 8:52.
Wei et al., 2012 (a) Brain IL-6 elevation causes neuronal circuitry imbalances and mediates autism-like behaviors, Biochimica et Biophysica Acta, 1822 (2012) 831–842.
Wei et al. 2012 (b) Alteration of brain volume in IL-6 overexpressing mice related to autism, International Journal of Developmental Neuroscience, 30:554-559.
Wei et al., 2013 Brain IL-6 and autism, Neuroscience 252 (2013): 320–325.
Wei et al., 2016 Inhibition of IL-6 trans-signaling in the brain increases sociability in the BTBR mouse model of autism, Biochimica et Biophysica Acta, 1862(10):1918-1925.
Weir et al., 2015 Preliminary evidence of neuropathology in nonhuman primates prenatally exposed to maternal immune activation, Brain, Behavior, and Immunity, 48,139–146.
Williams et al., 2006 The Profile of Memory Function in Children With Autism, Neuropsychology, 20(1): 21-29.
Wobke et al., 2014 Vitamin D in inflammatory diseases, Frontiers in Physiology, 5: 244.
Zerbo et al., 2014 Neonatal cytokines and chemokines and risk of Autism Spectrum Disorder: the Early Markers for Autism (EMA) study: a case-control study, Journal of Neuroinflammation, 11:113.
Zerbo et al., 2017 Association Between Influenza Infection and Vaccination During Pregnancy and Risk of Autism Spectrum Disorder, JAMA Pediatrics, 171(1).
Publikované: 18. augusta 2017 (verzia 1.0)
Akčná sieť pre informovaný súhlas

Anglický originál na stiahnutie tu:
https://www.icandecide.org/wp-content/uploads/2019/09/ICAN-AluminumAdjuvant-Autism.pdf

KONTAKT
PO BOX 18, 073 01 Sobrance
055 234 5793, 0907 534 623
info@pravonazivot.sk
![]()
![]()



